
Газета «Новости медицины и фармации» 6 (450) 2013
Вернуться к номеру
Система «теломеры — теломераза»: теория старения и пути к его замедлению — нутрицевтики. Актуальность проблемы
Авторы: Шушляпин О.И., к.м.н., врач-терапевт высшей категории; академик профильных академий ХНМУ: член Всемирной организации семейных врачей (WONCA), член Международной Петровской академии наук и искусств, CПб., РФ, член Нью-Йоркской академии наук, член Европейской ассоциации поддержки науки и технологий, Страсбург, Франция, член Лондонской дипломатической академии; Харьковский национальный медицинский университет, Украина
Рубрики: Семейная медицина/Терапия, Терапия, Геронтология
Разделы: Справочник специалиста
Версия для печати
 Многочисленные современные варианты первоначальной теории износа, возрастание энтропии, накопление генетических дефектов и повреждений, запрограммированность лимита существования (эффект Хейфлика), различного для каждого вида, смерть как вероятностное событие при взаимодействии с окружающей средой или как результат естественного отбора признаков (генов), обладающих одновременно полезными и повреждающими свойствами, — вот перечень ключевых воззрений на причины естественной смерти, возникающие на основе фундаментальных наук, в частности экологии, генетики, молекулярной биологии и онтогенеза [1, 2].
Многочисленные современные варианты первоначальной теории износа, возрастание энтропии, накопление генетических дефектов и повреждений, запрограммированность лимита существования (эффект Хейфлика), различного для каждого вида, смерть как вероятностное событие при взаимодействии с окружающей средой или как результат естественного отбора признаков (генов), обладающих одновременно полезными и повреждающими свойствами, — вот перечень ключевых воззрений на причины естественной смерти, возникающие на основе фундаментальных наук, в частности экологии, генетики, молекулярной биологии и онтогенеза [1, 2].
Теломерная теория старения и пути к продлению жизни основаны на изучении человеческой теломеразы, а соответственно, на попытках генетического омоложения человека.
Человечеству была бы нужна активная теломераза — «волшебный фермент молодости». Один из железных способов продлить жизнь — это принцип калорийноограниченного питания. Но эта пожизненная диета не ведет к увеличению средней и максимальной продолжительности жизни [3].
Теломеры — это маленькие участки на концах наших хромосом, и когда наши клетки делятся, ДНК в хромосомах должна копироваться, и во время этого процесса последняя маленькая частичка молекулярной цепи — кончик теломеры — не копируется, на протяжении нашей жизни теломеры постепенно укорачиваются. Это явление принято называть проблемой репликации концов хромосом, но которую следует назвать проблемой, запланированной заранее [4].
По мере укорочения теломер клетки стареют, хуже функционируют и реже начинают делиться, а стволовые клетки реже производят новые копии, а к какомуто моменту их совсем перестают производить. В результате этого человека подводит зрение, наша кожа теряет эластичность, хуже работает иммунная система и начинает происходить ряд других изменений, связанных со старением, и при этом нельзя изменить скорость возникновения главных болезней — патологии сердца и сосудов, злокачественных опухолей, цереброваскулярных болезней, гриппа и пневмонии (инфекционные болезни), атеросклероза, сахарного диабета, бронхита, бронхиальной астмы, нефрита, цирроза печени, если не удастся добиться замедления скорости старения. При этом старение — это своего рода болезнь, и задача геронтологии заключается не столько в продлении жизни, сколько в замедлении скорости старения, а значит, и прогрессирования главных болезней [5, 6].
Пути реализации проблемы или системные подходы исследований генов клеточного старения и долголетия человека c использованием нутрицевтиков
В настоящее время известно, что реактивация теломеразы продлевает репликативную жизнь соматических клеток, т.е. увеличивает число их делений. Однако это именно то, что происходит в опухолях и приводит их к злокачественному росту [7].
Как же достичь долголетия человека, замедлив старение, в то же время не вызывая развития злокачественных новообразований?
Одним из предлагаемых путей является реактивация теломеразы в пролиферирующих клетках на фоне стимулирования активности нормальных вариантов онкосупрессоров: р53, р16 и ARF. Так, было показано, что конститутивная сверхэкспрессия обратной транскриптазы (субъединицы теломеразы) с повышенной активностью ключевых супрессоров опухолей, перечисленных выше, приводит к значительному увеличению медианной продолжительности жизни и замедлению различных показателей старения, в частности кожи и кишечника [8].
Полагают, что клеточное старение — это генетическая программа необратимой остановки клеточного цикла, блокирующая реакцию клетки на пролиферирующие стимулы и факторы роста при наличии нерепарируемых повреждений ДНК, в частности критически укороченных телемер. Выдвигается даже гипотеза, что в эволюции клеточное старение возникло для того, чтобы не дать переродиться генетически поврежденной клетке в опухолевую, и определяющую роль в этом процессе, возможно, играет ген супрессора опухолей р53. Его продукт экспрессируется повсеместно во всех типах клеток в виде неактивного латентного транскрипционного фактора и активируется только тогда, когда клетка подвергается различным стрессам, таким как потеря теломер, повреждение ДНК, активация онкогенов или оксидативный стресс, и несмотря на то, что в стареющих клетках, в частности фибробластах, уровень белка р32 или его мРНК не увеличивается, но возрастает степень его фосфорилирования, а следовательно, возрастает ДНКсвязывающая активность, и в результате этого уровень основной мишени р53 и белка р21 в стареющих клетках остается значительно повышен и при этом нарастает с числом клеточных делений. Именно р21 отвечает за р53зависимую остановку клеточных делений. Р53зависимая индукция гена 21 приводит к клеточному старению на том основании, что белок р21 ингибирует клеточные регуляторы, в частности регуляторы клеточного цикла — циклинзависимые киназы, а также блокирует репликацию ДНК, связываясь с ядерным антигеном пролиферирующих клеток (PCNA), что и обусловливает необратимость клеточного цикла старения [9, 10].
Недавно было показано, что кодируемые другим локусом — INK4AFRF — белки p16 и р14 (ARF) также представляют собой клеточные регуляторы клеточного старения. Белок р16, как и р21, выступает в роли ингибитора циклинзависимых киназ. Экспрессия р16 заметно увеличивается с возрастом практически во всех тканях — это результат активации белка митогенактивируемой протеинкиназы р38 в ответ на оксидативный либо генотоксический стресс, которая есть не что иное, как опосредованное повышение уровня экспрессии р16 [11, 12].
Удивительным открытием было обнаружение того факта, что р53 играет определяющую роль при старении не только пролиферирующих клеток, но и постмитотических, таких как нейроны или клетки сердечной мышцы, и в этом процессе речь идет о р53зависимом апоптозе, когда, например, экспрессия доминантнонегативных, не способных связываться с ДНК, вариантов р53 в нейронах приводит к продлению на 10–20 % медианной и максимальной продолжительности жизни, но в то же время на других биологических образцах, например различных видах мышей с измененными сверхактивными формами р53, происходит процесс ускоренного старения, характеризуясь короткой продолжительностью жизни и ускоренным развитием возрастзависимых патологий, а при этом в ряде случаев добавочные копии гена р53, находящиеся под нормальным генетическим контролем экспрессии, характеризуются у предыдущих биологических объектов усиленным ответом на ДНК с низкой частотой возникновения злокачественной опухоли, но не проявляют признаков ускоренного старения или стареют медленнее [13].
Старение клетки — это загадочный феномен, возможно, играющий определенную роль в старении целостного организма, и как было показано у человека, при более низких концентрациях теломеразы и меньшей длине теломер в лимфоцитах периферической крови в несколько раз увеличивался риск сердечнососудистых заболеваний, составляющих, как известно, главную причину старческой смертности [14].
Программа клеточного старения может быть активирована в теломеразапозитивных клетках, каковыми являются стволовые клетки, и поэтому убыль количества стволовых клеток является одной из причин возрастзависимых нарушений регенерационной способности организма. Решение данной проблемы возможно на путях введения эмбриональных стволовых клеток в организм человека, но это может приводить к отторжению чужих клеток при их трансплантации. Перепрограммирование собственных дифференцированных клеток в подобие эмбриональных приводит к возникновению устойчивых свойств стволовых клеток благодаря обнаружению ряда регуляторных белков, обусловливающих свойства истинно стволовых клеток и увеличивающих продолжительность жизни человека [15].
Следующие факты указывают на то, что клеточное старение — репликативное и стрессиндуцированное, а также сокращение пролиферирующих клеток вообще и стволовых в частности приводит к нарушению регенераторной способности тканей, и на фоне возрастзависимой активации апоптоза в ряде тканей при подавлении компенсаторной пролиферации (замещение одних клеток другими) происходят дегенеративные нарушения, что снижает функциональные возможности организма, находя выражение в следующих процессах: длина теломер связана со старением целостного организма (1), модельные организмы с дефектами теломеразы характеризуются ускоренным старением и пониженной продолжительностью жизни (2), некоторые симптомы ускоренного старения у человека связаны с преждевременной дисфункцией теломер (3), стареющие клетки способны разрушать межклеточный матрикс, а также индуцировать локальное воспаление и онкогенез (4), снижение пролиферативных способностей или утеря стволовых клеток при старении снижает регенераторные способности любой ткани организма (5) [16].
Ведущие ученые мира видят перспективы исследований клеточного старения для решения новых задач в достижении качественного долголетия на пути:
1) разработки методов определения биологического возраста человека на основании специфических маркеров клеточного старения;
2) создания способов контроля экспресии генов, участвующих в клеточном старении, — теломеразы, р53, р21, р14 и ARF, фармакологическими и генноинженерными методами in vivo [17];
3) отработки безопасных подходов к перепрограммированию дифференцированных клеток в стволовые in vivo, в частности исследованиями, показавшими, что опосредованное ретровирусом введение четырех транскрипционных факторов (Oct4/Sox2/Klf4/cMyc) во взрослые дифференцированные клетки приводило к их переходу в клетки, что повышало продолжительность жизни человека [18].
На протяжении нескольких последних лет обсуждение возможности продления теломер сводилось главным образом к самому важному — к возможности включения этого гена, т.е. к активации теломеразы, и тогда полагали, что когда нам удастся это сделать, то тем самым станет возможным удлинять теломеры во всех клетках организма и таким образом со старением будет покончено. Однако последние исследования in vivo показали, что активация теломеразы не увеличивала среднюю длину теломера и даже не замедляла укорочение теломер в клетках [20].
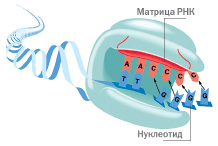
Теломераза — фермент, удлиняющий теломеры. Ген, который закодирован на это, присутствует в каждой клетке нашего организма. Однако в большинстве из клеток он не выражен, что означает, что он не превратился в теломеразу [19].
Все время, пока ведутся поиски по активации теломеразы, ученых мучает вопрос: что еще может замедлить укорочение теломер? Ответ оказался прост: «Нужно сохранять в целостности те теломеры, которые существуют в клетках, и принимать те натуральные средства или так называемые нутрицевтики, которые могут замедлить старение, воздействуя на его истинную причину — укорочение теломер», в противовес образному выражению «что имеем, то не ценим, потерявши — плачем» [21].
Одним из разработчиков данных по натуральным продуктам стала фирма под торговой маркой Terraternal, авторские права которой защищены международной лицензией на их продажу в 2008 году, а при создании продуктов по защите теломер были взяты результаты научных открытий. Благодаря соединению в одном мощном по воздействию продукте наиболее перспективные в этом отношении нутрицевтики — полезные для здоровья пищевые добавки при длительном курсовом их применении, которые можно заказать по адресу: http://www.terraternal.com/Products/ProductDetails/ru/TelomereGuard/616/155.aspx?gclid=CMLCsvv7obMCFY98c... [22].
Старение — разрушительный биологический процесс в соответствии с генетически запрограммированной теорией старения, описывается наиболее вероятный кандидат в гены старения, который называется «теломерa», при котором ограничивается приспособляемость организма и увеличивается вероятность смерти.
Старение является «дамокловым мечом», занесенным над каждым человеком начиная с 20летнего возраста. Возрастзависимые патологии: инфаркт, инсульт, онкологические заболевания, патология опорнодвигательного аппарата, психические и инфекционные заболевания пожилого возраста [23].
Заключение
Теломераза — фермент, который достраивает укороченную теломеру в половых клетках и клетках опухолей, обеспечивая их бессмертие на основе установленных следующих фактов:
1. Концы линейных хромосом с 3’конца ДНК заканчиваются повторяющимися последовательностями нуклеотидов, получивших название «теломеры», которые синтезируются рибонуклеиновым ферментом — теломеразой.
2. Соматические клетки эукариот, имеющие линейные хромосомы, лишены теломеразной активности. Их теломеры укорачиваются в процессе онтогенеза как по мере старения in vivo, так и при культивировании in vitro.
3. Половые клетки и клетки иммортализованных линий, а также опухоли имеют высокоактивную теломеразу, которая достраивает 3’конец ДНК, на котором реплицируется комплементарная цепь при делении.
4. Структуры теломер сильно различаются среди простейших, однако у всех позвоночных они одинаковы — (TTAGGG)n.
5. Имеются существенные межвидовые различия в длине теломер, причем у мыши общая их длина в несколько раз превышает таковую у человека (до 150 тыс. пар нуклеотидов у некоторых видов мышей и 7–15 тыс. пар нуклеотидов у человека).
6. Репрессия теломеразы характеризует клеточное старение в культуре (лимит Хейфлика).
7. Клетки больных синдромом преждевременного старения Хатчинсона — Гилфорда и синдромом Дауна имеют укороченные теломеры.
8. При введении теломеразы в клетки фибробластов человека, которые в норме делятся лишь 75–80 раз, способны поделиться 280 раз без какихлибо признаков старения и патологии. Тщательное исследование выявило, что в этих клетках нет таких признаков малигнизации, как, например, нестабильность хромосом, независимый от добавления натуральной сыворотки рост, отсутствие контактного торможения и потеря контроля над клеточным циклом. Кроме того, что особенно важно, из этих клеток не развиваются опухоли при трансплантации бестимусным мышам. Полученные данные свидетельствуют о том, что экспрессия теломеразы в культуре клеток человека совсем не обязательно вызывает развитие рака, т.е. теломераза лишена свойств онкогена. Основным свойством теломеразы является контроль клеточного деления, а для возникновения опухолевого роста необходимы дополнительные мутации и биохимические факторы.
9. Введение каталитического компонента теломеразы hTERT или теломеразной активности с помощью онкобелка вируса папиломы человека Е7 в кератоциты или в клетки эпителия человека не приводило к их полной иммортализации (смертельного исхода). Последняя наступала лишь при дополнительном торможении регуляции антионкогена Rb или при угнетении экспрессии р16 в качестве второй важнейшей ступени этого процесса. При элиминации антионкогена р53 такого эффекта не наблюдалось. С другой стороны, протоонкоген сМyc может активировать экспрессию теломеразы. С помощью опосредованного микроклетками переноса маркированную геном пео хромосому 20 из стареющих и молодых диплоидных фибробластов человека ввели в молодые фибробласты. Во всех новообразованных клонах наблюдалось уменьшение пролиферативного потенциала на 17–18 удвоений.
10. К восьмому десятку жизни частота деления кожи сокращается на 50 %.
Клиническое наблюдение
Размер и количество родинок связаны с длиной теломер, что представляет собой потенциальный маркер старения.
Родинки представляют собой один из важных факторов риска развития миелом. Они появляются в детстве и подростковом возрасте и подвергаются инволюции начиная со среднего возраста.
Изучалась длина теломер у 1897 женщин в возрасте от 18 до 79 лет. При этом количество родинок (невусов) положительно коррелировало с длиной теломер.
В среднем на теле взрослого человека можно найти от 30 до 100 родинок, однако в некоторых случаях их может быть более 400. Обнаружена статистически значимая корреляция по длине теломер между участниками исследования с менее чем 25 родинками на теле и теми, у которых их было более 100, в клетках которых теломеры были значительно длинней. Это означало, что по «теломерному маркеру» клетки этих людей были моложе приблизительно на 6–7 лет.
Отсюда вывод: понимание механизмов влияния индукции и инволюции родимых пятен не только поможет в понимании патофизиологии меланомы, но также должно пролить свет на сложную связь между старением и раковыми опухолями. Кроме того, незначительный риск злокачественных новообразований, связанный с большим количеством родинок, с лихвой компенсируется меньшей склонностью к возрастным заболеваниям.
1. Акоев И.Г. Биофизика познает рак. — Москва: Наука, 1987. — 160.
2. Милунски О. Знайте свои гены. — Москва: Мир, 1881. — 392.
3. Каминер Т., Гильхрест В.А. и др. Старение кожи // Principles of Geriatric Medicine and Gerontology. — New York: McGrawHill, 1994. — Вып. 3. — 411429 s.
4. Яар М., Гильхрест В.А. Старение кожи: теоретические допустимые механизмы и вытекающие из них изменения в структуре и функционировании // Clin. Geriatr. Med. — 2001. — 17. — 617630.
5. Дильман В.М. Четыре модели медицины. — Ленинград: Медицина, 1987. — 288.
6. Дильман В.М. Большие биологические часы. — Москва: Знание, 1982. — 208.
7. Фанк В.Д., Ванг С.К., Шелтон Д.Н., Шарли С.В. и др. Экспрессия теломеразы восстанавливает целостность кожи у состарившихся in vitro фибробластов в воссозданной модели кожи // Exp. Cell. Res. — 2000. — 258, 270278.
8. Егоров Н.С., Олексин А.В., Самуилова В.Д. Биотехнология: Учебное издательство для вузов. Книга 1: Проблемы и перспективы. — Москва: Высшая школа, 1987. — 159.
9. Грунченко Е.В. Что нам стоит многоклеточность. — Новосибирск: Наука, 1985 (сибирское отделение). — 135.
10. Сарецки Г., фон Цглиники. Репликационное старение, теломеры и оксидативный стресс // Ann. N. Acad. Sci. — 2002. — 959. — 2429.
11. Эффрос Р.Б. Старение и иммунная система. Восприимчивость и старение: причины и вмешательство / Novartis Foundation Symposium; еds G. Bock and J.A. Goods. — John Willey & Son, Ltd., Chicherst, UK, 2008. — 235.
12. Лоренц М., Сарецки Г., Ситте Н. и др. Фибробласты ВJ демонстрируют высокие оксидативные возможности и замедляют укорачивание теломер независимо от hTERTтрансферазы // Free Radic. Biol. Med. — 2001. — 31. — 824831.
13. Фаyce, Эфрос. Фармакологическое усиление антивирусной функции CDB+Тлимфоцитов на основе теломеразы // Immunology. — 2008, Nov. 15. — 181(10). — 74007406.
14. Канунго М. Биохимия старения. — Москва: Мир, 1982. — 296.
15. Лэмб М. Биология старения. — Москва: Мир, 1980. — 206.
16. Йоку, Фурумото и др. Связанное со старением укорачивание теломер замедляется обогащением внутриклеточного витамина С путем подавления оксидазного стресса // Life Sci. — 1996. — 63(11). — 935938.
17. Ричард Г.А., Фарагер, Дэвид, Киплинг. Как репликативное старение может влиять на старение человека // Bioеssay. — 1998, Des. — Т. 20, вып. 12. — 985991.
18. Гуо Н., Перри Э.М., Ли Л.С., Кэмбу Ф., Лодер Н. и др. Короткие теломеры подвергают риску передачу сигналов и долговечность bклетки // Nature. — 2011. — 6(3), PloS ONE. — e17858.
19. Шей, Райт, Баур. Влияние положения теломеры в клетках человека // Sciense. — 2001. — Т. 292, № 5524. — 20752077.
20. Эргюнм, Сахин и др. Дисфункция теломеров вызывает метаболическую митохондриальную компроментацию // Nature. — 2012. — 470. — 359365.
21. Бласко и др. Активатор теломеразы ТА85 удлиняет короткие теломеры и увеличивает продолжительность жизни взрослых/старых мышей, не увеличивая частоту заболевания раком // Aging Cell. — 2011. — Т. 10, вып. 4. — 604621.
22. Харли, Луи, Бласко и др. Натуральный продукт — активатор теломеразы как часть программы сохранения здоровья // Rejuvenation. — 2010. — Т. 14, № 1. — 3439.
23. Майк Фоссель. Клетки, старение и заболевание человека. — Oxford University Press, 2004. — 290.